![]()
visites sur cette page
|
|
Recherches personnelles principales |
Le Dévonien |
||
|
Avant le Dévonien
|
||||
avec le concours de Frédéric Boulvain (ULG), Léon Dejonghe (ULB), Johan Yans (UNamur) et Alain Préat (ULB)
Qu'ils soient remerciés pour leurs conseils éclairés, leurs suggestions de visites sur le terrain, les lieux de prospections intéressants et la mise à ma disposition de certaines publications spécifiques.Plan de la page
Situation du Dévonien dans l'échelle géologique
Géologie du Dévonien
Introduction
Chronologie des formations et histoire de la région
Les tenants et les aboutissants : avant le Dévonien...
Le CambrienLe Dévonien...
L'Ordovicien
Les premières communautés terrestres : des écosystèmes dynamiques
Un passé mal connuLa grande extinction qui marque la fin de l'Ordovicien
Conclusion
Les ancêtres des Embryophytes
La première communauté complexe et bien documentée : Rhynie
Le Silurien
Introduction à la GéologieLe Dévonien Inférieur
Le LochkovienLe Dévonien Moyen
Le Praguien
L'Emsien
L'EifelienLe Dévonien Supérieur
Le Givetien
Le FrasnienL'explosion des végétaux
Le Famennien
L'avènement de la vie terrestre
La fin du Dévonien
Une approche du monde végétal.De l'arbre à la forêt : Influence d'une innovation végétale dévonienne sur les interactions biosphère / atmosphère / hydrosphère.
La photosynthèse.Les tendances évolutives des végétaux.
La différenciation des végétaux.
Qu'est-ce qu'un arbre?
Le temps des bactéries et des algues.
La conquête des terres émergées.
Les conséquences de l'implantation des végétaux
Les plantes pionnières, cause de la glaciation ordovicienne ?
La diversification du monde végétal au Dévonien.L'apparition des herbivores et leur influence sur l'évolution des écosystèmes.
Les premiers arbres
Le développement des forêts modifie la planète.
Les forêts du Dévonien supérieur au Permien.
Le temps des fougères
Quelques rappels sur le fonctionnement des écosystèmes.
Les arthropodes herbivores : apparition et impact sur la dynamique des écosystèmes
Les premiers vertébrés herbivores apparaissent dans différents groupes.
Bilan sur l’évolution des interactions animaux / végétaux.
Le temps des Gymnospermes.
Conclusion.
Le Dévonien constitue une période de transition entre les deux grands cycles orogéniques calédonien et hercynien. Sur le plan paléontologique, rappelons que le Dévonien correspond à un tournant fondamental de l'évolution de la vie : la conquête du monde continental.
Voici ci-dessous la situation du dévonien dans l'échelle géologique. Avant de s'intéresser proprement dit au Dévonien, il serait intéressant de situer cette période dans l'échelle géologique afin d'en avoir une idée générale.
Millions
d'annéesEonothème Erathème
Système
Epoque
Etage
Principaux évènements
N° 0,0012
PHANEROZOIQUE
Quaternaire supérieur
Holocène supérieur
Néolithique
Début de l'âge du fer
12500 0,0015
Installation de civilisation proto-celtes en Gaule
12400 0,0016
Début de l'âge du bronze
12300 0,0018
Installation les Ligures en Europe Occidentale
12200 0,0025
Début de l'âge du cuivre
12100 0,0075
Sédentarisation
apparition de l'élevage, de l'agriculture, de la poterie, du tissage et des premiers mégalithes
12000 0,015
Mésolithique
11900 0,020
Holocène inférieur
Paléolithique
Fin de la glaciation de Würm
11800 0,025
Disparition de l'homme de Neandertal
11700 0,035
Début de la civilisation Moustérienne du Châtel
11600 0,04
Arrivée de l'Homo Sapiens en Europe
11500 0,05
Apparition des premières peintures rupestres
11400 0,07
Début de la civilisation Moustérienne
11300 0,08
Début de la glaciation de Würm
Premières sépultures
11200 0,13
Fin de la glaciation de Riss
11200 0,30
Début de la glaciation de Riss
11100 0,20
Apparition des premiers Homo Sapiens
11000 0,40
Début de la glaciation de Mindel, domestication du feu
10900 0,65
Début de la période interglaciaire Günz-Mindel
10800 0,80
Quaternaire inférieur
Pléistocène
Supérieur
Apparition des premiers Hommes de Neandertal en Espagne (Atapuerca)
10700 0,95
Début du pré-paléolithique
Début de la glaciation de Günz
10600 1,2
Apparition de l'Homo Erectus dans le Sud de la France
Disparition de l'Australopithecus Robustus
10500 1,3
Disparition de l'Homo Habilis
10400 1,6
Inférieur
Apparition de l'Homo Erectus
10300 1,8
Première présence d'outils (galets) en France en Haute-Loire
Apparition des premiers outils symétriques (bifaces) près du lac Turkana
10200 2
Première présence de l'Australopithecus Robustus
10100 2,5
CENOZOIQUE
(Tertiaire)
Néogène
Pliocène
Gélacien
Apparition de l'Homo Habilis et des premiers outils
10000
3
Première présence de l'Australopithecus Africanus
3,18
Présence de l'Australopithecus Afarensis (Lucy) près de Hadar
3,5
Plaisancien Empreintes de préhumains près de Laetoli
9900 3,7
Première présence de l'Australopithecus Afarensis près de Laetoli
4
Zancléen
Apparition de l'Australopithecus Anamensis
9800 4,4
Présence de l'Australipothecus Ramidus dans la vallée de Lawash
5,5
Miocène
Messinien
Présence d'Australopithèques près du Lac Turkana
9700 6
Premiers Australopithèques près de Lothagam
Millenium Ancestor, ancêtre probable de l'homme moderne
8
Tortonien
Séparation de la lignée des Primates et de Hominidés, début de l'East Side Story
9600 15
Serravalien
Apparition du Kenyapithèque
9500 20
Langhien
Apparition du Proconsul
9400 22
Burdigalien
Séparation de l'Australie de l'Antarctique
9300 23
Aquitanien
9200 29
Paléogène
Oligocène
Chattien
9100 35
Rupélien
Apparition des Rhinocéridés
9000 37
Eocène
Priabonien
8900 42 Bartonien 8800 49
Lutétien
Cerithium giganteum
8700 56
Yprésien
Apparition des premiers Primates
Explosion des Mammifères
8600 60
Paléocène
Thanétien
Epanouissement des Mammifères
Apparition des Insectivores
8500 64
Dano-Montien
Explosion des plantes à fleurs
8400 65
MESOZOIQUE
(Secondaire)
Crétacé
Supérieur
Maastrichtien
Chute d'une météorite géante
Fin des
Dinosauriens
et
des AmmonitesApparition des Primates
Formation de l'Atlantique Nord
Apparition des premières plantes à fleurs
8300 72
Campanien
8200 83
Santonien
8100 85
Coniacien
8000 88
Turonien
7900 95
Cénomanien
7800 107
Inférieur
Albien
Formation
de l'Atlantique Sud
Apparition des oiseaux et marsupiaux
7700 110
Aptien
7600 112
Barrémien
7500 114
Hauterivien
7400 119
Valanginien
7300 125
Berriasien
7200 130
Jurassique
Malm
Portlandien
Explosion des ammonites
7100 140
Kimméridgien
7000 145
Oxfordien
6900
150
Dogger
Callovien
6800 160
Bathonien
6700 167
Bajocien
6600 176
Aalénien
6500 181
Lias
Toarcien
Apparition de la famille des palmiers
Apparition des dinosaures aériens et marins
6400 188
Pliensbachien
6300 195
Sinémurien
6200 204
Hettangien
6100
212
Trias
Réthien Fin de la Pangée
Premiers Mammifères
6000 220
Norien
5900 230
Carnien 5800 235
Ladinien 5700
240
Anisien 5600 245
Scythien 5500 250
PALEOZOÏQUE
(Primaire)
Permien
Supérieur Thuringien
Premiers Dinosaures
5400 270
Inférieur Saxonien
Glaciation 5ème extinction massive des végétaux et des animaux
5300 290
Autunien
Conifères
5200 300
Carbonifère
Silésien
(Houiller)
Stéphanien
Reptiles
5100 310
Westphalien
Fougères arborescentes
5000 320
Namurien
Insectes
4900 340
Dinantien Viséen
Amphibiens
4800 360
Tournaisien
Fougères
Poissons
osseux4700 367
Dévonien
Supérieur
Famennien
Plantes
terrestresPremiers tétrapodes
4600 375
Frasnien
Glaciation
4ème extinction
Premières ammonites
4500 378
Moyen Givetien
Stringocephalus burtuni
4400 382
Eifelien
Calceola sandalina
4300 385
Inférieur Emsien
4200 387
Praguien
Paraspirifer cultrijugatus
4100 390
Lochkovien
Chlorodictum problematicyum
4000 410
Pridolien
Schistes Bigarrés d'Oignies et de Saint Hubert
3900 415
Silurien Ludlowien Ludfordien
Sortie de l'eau des plantes vasculaires
3800
Gorstien
Actinoptérygiens
3700 425
Wenlockien
Homérien
Ostéichtyens
Myriapodes
3600 430
Scheinwoodien
Gnathostomes
3500
Llandovérien Telychien
3400 435
Aeronien
Arachnides
3300 440
Rhuddanien
Sortie de l'eau des algues
3200 445
Ordovicien
Supérieur
Ashgillien
3100
Caradocien
Apparition des premiers poissons et des mollusques céphalopodes
3000 455
Inférieur
Llandeilien
Trilobites
2900
Llanvirnien
2800 470
Arénigien
Premiers végétaux terrestres
2700 485
Trémadocien
2600 500
Cambrien
Postdamien
Trempoéaléouien
Apparition des éponges, des mollusques, des trilobites, des échinodermes
2500
Franconien
2400 Dresbachien 2300 Acadien Mayaien 2200 Amgaien 2100 Lénien 2000 530 Géorgien Atdabatien 1900 Tommotien 1800 540 Némakit-Daldynien 1700 630 PROTEROZOÏQUE Néoprotérozoïque
Ediacarien Faune d'Ediacaria 1600 850 Cryogénien Apparition des premiers vers
Glaciation de Sturtien
Glaciation de Varangien
1500 1000 Tonien Fragmentation de Rodinia Début du règne des Acritarches (algues vertes)
1400 1200 Mésoprotérozoïque
Sténien Glaciation Formation du super continent Rodinia
1300 1400 Ectasien Algues rouges Bangiomorpha pubescens 1200 1600 Calymnien Apparition des premiers métazoaires (êtres pluricellulaires) Fragmentation de Columbia
1100 1800 Stathérien 2ème extinction
Formation de plateaux continentaux
Continent Columbia
1000 2050 Paléoprotérozoïque
Orosirien Apparition des bactéries coccoïdes (ancêtres du phytoplancton)
Oxygénation de l'atmosphère
Orogenèse sur les terres émergées
900 2300 Rhyacien Glaciation huronienne
Groupe fossile de Franceville
800 2500 Sidérien Formation de fer rubané 700 2800 ARCHEEN
Néoachéen Apparition des Eucaryotes (cellules présentant un noyau) 600 3200 Mésoarchéen Apparition des algues bleues et de la photosynthèse 500 3400 Paléoarchéen Développement des stromatolites
Impact d'une météorite géante
(Fig Tree)
1ère extinction
400 3800 Eoarchéen Apparition des premières formes de vie (bactéries)
300 4500 HADEEN ETOILES ET PLANETES Naissance du système solaire 200 15000 BIG BANG Naissance de l'Univers 100
Détaillons l'échelle des temps géologiques ci-dessus et présentons les différentes couches observables avec positionnement dans le temps par rapport aux autres couches des autres étages, systèmes et groupes.
Sur mon échelle géologique personnelle, j'ai numéroté les différentes couches de 100 en 10 en partant du Big Bang (100) jusqu'à l'Actuel (12500). Ce travail un rien fastidieux me permet et vous permettra de situer plus facilement les différents affleurements que nous rencontrerons. Voir Echelle géologique
Dans le Dévonien, nous observerons des roches depuis le Poudingue de Fépin jusqu'à l'Assise de Comblain au Pont.
Voir Echelle géologique
Le nom "Dévonien" dérive du comté du Devonshire (Sud-Ouest de l'Angleterre). Le système est subdivisé en 5 étages dont chacun est subdivisé en sous-étages, en formations, en membres, en assises et enfin en couches.
Les subdivisions sont différentes selon que l'on considère les Ardennes, l'Angleterre ou l'Amérique. En ce qui me concerne, je me bornerai à la nomenclature belge et française de manière à pouvoir garder et employer les appellations "Famennien", "Frasnien", "Givetien", "Eifelien" (ancien Couvinien) et "Emsien", couches faisant référence respectivement à la Famenne, à Frasnes-lez-Couvin, à Givet et à Couvin selon les anciennes appellations, nous resterons cantonnés dans notre région : La Calestienne.
Le cycle varisque (anciennement appelé hercynien) s'est déroulé au Paléozoïque supérieur. A l'échelle des masses continentales, l'orogenèse calédonienne a soudé le continent Nordique Euramerica, encore séparé de Siberia et de Gondwana par un domaine océanique. Notre région est située sur la marge Sud de l'Euramerica, aussi appelé "Continent des Vieux Grès Rouges", en raison du dépôt de vastes séries détritiques à faciès continental résultant de l'érosion de la chaîne calédonienne.
Le Dévonien constitue une période de transition entre les deux grands cycles orogéniques calédonien et varisque. Une certaine relaxation tectonique y est observée. Les faciès sont à dominance détritique, ce qui indique que l'érection des montagnes s'arrête et que l'érosion des massifs prend le dessus, amenant per le biais des rivières et des fleuves, des sédiments qui se déposent dans les océans. Au Carbonifère inférieur, une transgression générale, associée à une diminution des apports détritiques, permet le développement de vastes plates-formes carbonatées, tandis que commencent à émerger les premiers reliefs des chaînes varisques, alimentant localement une sédimentation de type flysch (faciès Kulm). Au Carbonifère moyen et supérieur, les dépôts houillers vont s'accumuler en bordure de la chaîne varisque en formation d'abord, dans des fossés d'effondrement ensuite.
Au Permien, les masses continentales sont réunies en une Pangée, vaste continent unique au climat aride.Sur le plan paléontologique, rappelons que le Paléozoïque supérieur est marqué par la conquête des continents : les végétaux vont envahir le milieu terrestre jusqu'à constituer d'immenses forêts au Carbonifère. Ceci entraînera des modifications fondamentales à l'échelle de la planète, notamment dans la diminution du CO2atmosphérique et dans l'altération continentale, nettement contrôlée par la couverture végétale. Les animaux également conquièrent le milieu aérien : on observe l'apparition des batraciens et des reptiles ainsi que des premiers insectes. Sur le plan bio stratigraphique, les groupes les plus importants sont les goniatites, les trilobites, les fusulines et les végétaux.
Mais resituons géologiquement le Dévonien au sein de la paléoécologie : formation des roches, dérive des continents, érection de montagnes, transgressions marines, régressions marines, érosions, règnes animaux et végétaux se succédant, luminosité, chaleur, humidité, climat...
Racontons l'histoire de cette région et tentons d'y inclure les différentes couches observées, ce qui nous permettra d'expliquer leurs formations.
Chronologie des formations et histoire de la région
Les tenants et les aboutissants : avant le Dévonien...
L'histoire de ces roches a commencé il y a 500 à 600 millions d'années, au sein d'une mer peu profonde qui recouvrait notre région. Quand nous disons "notre région", il faut comprendre en fait une région située dans l'hémisphère Sud, aux environs de 70° de latitude Sud.
Position des continents il y a 600 millions d'années, soit à la toute fin du Précambrien, alors que ceux-ci dérivaient les uns par rapport aux autres : Laurentia, Baltica, Sibéria, Gondwana, Kazakhstania, China. Progressivement, s'ouvrait un océan entre Laurentia et Baltica, un océan que les géologues ont baptisé Iapétus.
Il faut se rappeler que l'écorce terrestre n'est qu'une mince pellicule au regard de notre planète. Cette fine peau est scindée en plaques qui dérivent très lentement les unes par rapport aux autres. Depuis le Cambrien, le socle de notre pays, situé alors au Sud de l'équateur a dérivé lentement vers le Nord pour atteindre aujourd'hui nos latitudes.
Début du Cambrien, il y a 540 millions. L'océan Iapetus s'ouvrait encore. On doit donc supposer l'existence d'une dorsale médio-océanique entre Laurentia et Baltica. Gondwana amorçait une migration vers le Sud.
Au cours du Cambrien, des sables, fins, argileux, se sont déposés au fond de la mer. Ils sont à l'origine des quartzites. Alternativement, les dépôts sableux laissaient la place à des boues fines qui furent à l'origine des schistes et des phyllades. La teinte noire des roches est due, en partie, à des sulfures de fer, qui, par exposition à l'air se sont altérés en oxydes de fer. A l'origine, les dépôts successifs de boues et de sables fins se sont superposés en couches parallèles presque horizontales. Cette superposition se retrouve ici dans la région de Chimay, Momignies, Cul-des-Sart... mais les bancs s'inclinent vers le Sud-est.
Au milieu du Cambrien, il y a 525 millions d'années, Laurentia et Baltica s'étaient éloignés encore plus l'un de l'autre, produisant un océan Iapetus encore plus large. Gondwana poursuivait sa migration vers le Sud.
Vers la fin du Cambrien, il y a 510 millions d'années, l'océan Iapetus avait atteint son ouverture maximale après 140 millions d'années (flèche rouge à double pointe).
-524Ma Cambrien
Postdamien
Trempoéaléouien Apparition des éponges, des mollusques, des trilobites, des échinodermes
Franconien Dresbachien -545Ma Acadien Mayaien Amgaien -560Ma Lénien Géorgien Atdabatien Cette position des couches de schistes et de quartzites est due aux plissements qui ont suivi le Cambrien. Au cours de l'Ordovicien et du Silurien (entre 500 et 400 millions d'années avant notre époque actuelle), les lents mouvements de l'écorce terrestre ont relevé le fond marin jusque la surface. Cette plaine a continué à se soulever pour former un plateau de plus en plus élevé.
Dessin numérique original L.V.B.
Ces mouvements étaient très lents (de l'ordre du cm par siècle, au plus du mm par an) mais sur des millions d'années, la dérive des continents a provoqué des pressions latérales qui ont bousculé et plissé les couches de sédiments. Le continent ainsi formé va être la proie de l'érosion. Notons aussi pour être complet que ces boues argileuses et ces sables qui étaient au départ des roches meubles sont devenues d'une part suite à la compression des couches qui s'accumulent les unes sur les autres et d'autre part suite à la compression latérale due aux divers plissements, sont devenues, donc, des roches compactes et dures : les schistes et les quartzites.
-445Ma Ordovicien Supérieur Ashgillien Caradocien Apparition des premiers poissons et des
mollusques céphalopodes-455Ma Inférieur Llandeilien Trilobites Llanvirnien -470Ma Arénigien Premiers végétaux terrestres -485Ma Trémadocien Nous savons que le niveau de la mer est élevé pendant l’Ordovicien, la transgression marine du Trémadocien est celle où le niveau de la mer est le plus haut dont il reste des traces.
A la fin du Cambrien (vers -500 millions d'années), la région d'Europe Occidentale qui nous intéresse, se situait aux environs de 65° de latitude Sud et les terres qui la composaient formaient le "Gondwana" (Amérique du Sud, Afrique, Australie, Antarctique, Europe du Sud, Asie du Sud et Floride).
Au début de l’Ordovicien, Gondwana est situé aux latitudes équatoriales et se déplace vers le pôle Sud.
Au milieu de l'Ordovicien, alors que les terres continuent à dériver, de petits terranes se séparent du Gondwana. Cette séparation produit quelques petites plaques continentales qui dérivent alors de manière indépendante les unes des autres. Elles seront ré-accrétées au bloc principal durant l’Ordovicien moyen. Ces évènements orogéniques ont laissé des traces comme la présence en Europe d’ophiolites, de granites et de roches métamorphiques de cet âge (massifs cristallins des Alpes : aiguilles Rouges, Belledonne). Une de ces plaques s'appelait "Brabantia". Elle comprenait le Sud de l'Irlande, le Sud de l'Angleterre, la Belgique, les Pays-Bas et le Nord-Ouest de l'Allemagne.
Il y a 500 millions d'années, au tout début de l'Ordovicien, soit 150 millions d'années après le début de l'ouverture de Iapetus, il s'est développé à la marge de Laurentia une zone de subduction, créant du même coup un arc volcanique insulaire. Le mouvement s'était renversé. L'océan Iapetus commençait à se refermer. Laurentia et Baltica convergeaient. On est donc passé d'un océan de type Atlantique, c'est-à-dire en ouverture avec marges passives, à un océan de type Pacifique, en fermeture, avec marges actives. Durant tout ce temps, Gondwana migrait toujours vers le Sud.
Tout comme l’Amérique du Nord et l’Europe, Gondwana est en partie couvert de mers peu profondes. Les roches de l’Ordovicien sont principalement sédimentaires, les sédiments marins qui en forment la plus grande partie sont constitués en grande partie de calcaire; les schistes et le grès sont bien moins fréquents.
L’orogenèse taconienne se produit lors de la collision de Baltica et de Laurentia. Vers la fin de cette période, le Gondwana était proche du pôle et largement recouvert de glaciers, le climat se refroidit alors que la biodiversité croît.

Dessin L.V.B.
La grande bio diversification ordovicienne ou radiation ordovicienne est la période de l'histoire de la Terre pendant laquelle la biodiversité de la vie océanique a le plus augmenté. Après l'explosion cambrienne correspondant à l'éclosion de la plupart des phylums animaux, la radiation ordovicienne s'achève sur une remarquable diversité des niveaux inférieurs de la classification des taxons : ordres, familles, genres et espèces.
Cette phase de diversification biologique intervient environ à 60 millions d'années après l’explosion cambrienne. Sa durée est de l’ordre de 25 millions d'années (intervalle relativement court à l'échelle des temps géologiques), et se situe durant la partie inférieure et moyenne du système Ordovicien, datée entre 485 et 460 millions d'années.
Cette radiation majeure appelée en anglais the Great Ordovician Biodiversification Event (GOBE), porte aussi parfois le nom d'explosion ordovicienne. La radiation ordovicienne a été évaluée de façon quantitative par les paléontologues en se basant sur le dénombrement et l'évolution du nombre des taxons au Paléozoïque. Le nombre de familles d'animaux marins recensés va augmenter considérablement, passant d'une centaine à plus de 400 durant la radiation ordovicienne. Le nombre de genres évolue d'environ 500 à plus de 1 600.
Cet événement concerne la plupart des groupes d'animaux marins : brachiopodes, échinodermes, coraux, mollusques, graptoloïdes, conodontes, acritarches, etc. Il en est de même pour le micro phytoplancton qui voit son nombre d'espèces croître d'environ 200 à 500.
La découverte de plusieurs gisements d'âge Ordovicien inférieur dans la formation des argiles de Fezouata, dans la région de Zagora (Anti-Atlas central au Maroc), a montré la persistance des faunes du Cambrien sur une durée bien plus longue qu'envisagé initialement. Ces faunes rappellent celles du célèbre site des schistes de Burgess au Canada avec des animaux caractéristiques (anomalocarides, marrellomorphes, etc.) qui cependant coexistent avec des taxons typiquement ordoviciens (cirripèdes, euryptérides, limules). La transition entre l'explosion cambrienne et grande bio diversification ordovicienne s'avère moins brutale que précédemment décrit.


A gauche : fossiles d'Ampyx priscus du biotope de Fezouata (Trémadocien du Maroc) alignés en «file indienne» lors d'une migration ou d'un regroupement.
Au centre : fossile du trilobite Parapilekia cf. olesnaensis du biotope de Fezouata (Tremadocien du Maroc) conservé au Musee de Huston.
A droite : fossile d'un tout petit marrellomorphe Furca mauritanica du biotope de Fezouata (Trémadocien du Maroc)parfaitement conservé. (Muséum de Toulouse).Les causes du phénomène sont difficiles à cerner, d'autant que l'événement est ancien. De nombreuses études sont toujours en cours.
À l'instar des extinctions massives qui ont marqué l’histoire de la Terre, un grand nombre de causes sont aujourd'hui envisagées pour expliquer la radiation ordovicienne. Une combinaison de facteurs extrinsèques (géologiques) et intrinsèques (biologiques) est probablement à l'origine de ces phénomènes de bio diversification et de radiation. Les causes d’apparition, de modification, de préservation ou de disparition des habitats marins de cette époque sont étudiées en utilisant toute la palette des techniques de la géologie, dont la paléontologie, la sédimentologie, la paléogéographie, la paléoclimatologie, les analyses isotopiques, la géochimie, etc.
Parmi les causes possibles :
La configuration paléogéographique particulière de cette période. Au cours de l’érathème Paléozoïque, c’est en effet à l’Ordovicien que les masses continentales sont les plus dispersées du fait de la tectonique des plaques. Ces conditions favorisent la diversité des environnements et des habitats.
- L’Ordovicien inférieur et moyen est une période d’intense expansion océanique à partir des dorsales, qui coïncide avec le niveau marin le plus élevé du Paléozoïque (qui va atteindre avant la fin de l'Ordovicien une cote plus de 200 mètres plus haute que le niveau actuel des océans. Les eaux marines transgressives sur les continents forment des plateformes continentales, peu profondes, d’une étendue exceptionnelle. C'est durant cet intervalle que les plateformes intertropicales atteignent leur plus grande superficie au cours des temps géologiques. Ces environnements sont connus comme les plus propices à la diversification des faunes marines.
- L’intense activité volcanique de cette période favorise la disponibilité d’éléments nutritifs dans les océans qui permettent le développement considérable du phyto- et zooplancton, qui eux-mêmes stimulent le développement d'organismes qui s'en nourrissent, induisant ainsi un profond changement des chaînes alimentaires.
Malgré une baisse sensible de la température des océans (qui devait être supérieure à 40°C à la base de l'Ordovicien), le climat est encore chaud, probablement équivalent à celui des zones équatoriales actuelles. Ce niveau de température propice serait un des moteurs de la bio diversification.
Enfin, le rôle d'une importante «pluie météoritique» issue de la fragmentation d’un énorme astéroïde a été envisagé. Les multiples cratères d'impact auraient pu, à la fois détruire des environnements, mais aussi créer de nouveaux habitats prêts à être colonisés. Les traces de l'événement sont enregistrées dans les sédiments par la présence de grains de chromite extra-terrestre mise en évidence aussi bien en Scandinavie qu'en Chine. La datation de ces impacts à 470 millions d'années, légèrement décalée par rapport au début de la radiation ordovicienne, fait que le rôle de ces fragments d'astéroïdes est controversé.
Il y a 480 millions d'années, la fermeture de Iapetus se poursuivait et les arcs volcaniques insulaires fonctionnaient toujours.
Le rapprochement des masses continentales ne se faisait pas uniquement entre Laurentia et Baltica, mais aussi entre Laurentia et Siberia. Au Sud, une petite masse continentale s'était détachée de Gondwana et migrait vers le Nord.
Au milieu de l'Ordovicien, il y a 460 millions d'années, Iapetus continuait à se refermer. L'arc volcanique insulaire qui se trouvait au large de Laurentia, entra en collision avec la marge de Laurentia. C'est une collision de type plaque océanique contre plaque continentale : une chaîne de montagne immature s'est formée ; on a appelé cette chaîne, la chaîne taconienne, la première phase de la formation des Appalaches.
Le continent Gondwana a atteint le pôle Sud.
A la fin de l'Ordovicien, il y a 440 millions d'années, l'espace océanique entre Laurentia, Baltica et Siberia continuait à se refermer. La petite masse qui s'était détachée de Gondwana et qui migrait vers le Nord s'est morcelée pour donner un chaînon de microcontinents. Le pôle Sud était occupé par la marge de Gondwana, plus spécifiquement le Nord de l'Afrique actuelle. Signalons ici qu'on connaît au Maroc des dépôts glaciaires d'âge Ordovicien supérieur. Ce n'est pas surprenant, puisque le Nord de l'Afrique se situait au pôle Sud à cette époque.
L'extinction Ordovicien-Silurien est la disparition d'une proportion significative des espèces vivant sur la Terre (essentiellement marines à cette époque), il y a environ 445 millions d'années, à la limite entre l'Ordovicien et le Silurien.
L'extinction Ordovicien-Silurien, appelée parfois «extinction ordovicienne», est considérée comme la seconde plus importante des cinq grandes extinctions massives de l'éonothème Phanérozoïque. Elle aboutit à la disparition de 27% des familles et de 57% des genres d'animaux marins et une estimation de 85% au niveau des espèces.
La cause principale de cette extinction majeure paraît liée à une grande phase de volcanisme combinée avec une glaciation à la fin de l'Ordovicien. Cette glaciation aurait entraîné des désordres climatiques et écologiques rendant difficile l'adaptation des espèces et écosystèmes au recul de la mer sur des centaines de kilomètres, puis à son retour en fin de phase glaciaire.
Comme pour la plupart des extinctions massives un certain nombre de causes, parfois concomitantes et dépendantes, sont invoquées (tectonique des plaques, variations climatiques et écologiques, volcanisme, impacts d'astéroïdes, sursaut gamma, cycles astronomiques, concentration élevée en métaux lourds, etc.). Les scientifiques s'accordent sur les rôles majeurs de la grande glaciation qui a sévi à la fin de la période ordovicienne.
Au cours du Cambrien et durant la majeure partie de l'Ordovicien les masses continentales se sont largement dispersées favorisant une radiation évolutive remarquable : la Grande bio diversification ordovicienne. Cependant, à l'Ordovicien supérieur, certains de ces blocs continentaux entament un rapprochement qui conduira, au cours du Silurien, à la formation du supercontinent "Laurussia". L'autre supercontinent, le Gondwana dérive vers le Sud et, à la fin de l'Ordovicien, il se retrouve sous de très hautes latitudes australes. Il va alors se couvrir d'une épaisse calotte polaire entourée d'une large banquise. Cette glaciation, qui était considérée lors d'études antérieures comme un événement relativement bref, est en fait intervenue sur une période plus longue (plus de 10 millions d’années).
C'est lors de l'Ordovicien supérieur que les plantes non vasculaires s'installent et se développent sur la terre ferme. Cette modification majeure de la biosphère aurait accéléré le processus d'altération des silicates sur les continents. Ce processus, en fixant de très grandes quantités de dioxyde de carbone d’où une diminution drastique des gaz à effet de serre qui aurait conduit à la baisse des températures de la Terre et au développement des calottes polaires.
Les conséquences de cette glaciation sont multiples :
La conséquence majeure est la baisse du niveau de la mer qui entraine une réduction considérable des environnements de plateforme continentale et par conséquent de leurs habitats et de la biodiversité marine.
- Les bassins sont souvent isolés les uns des autres ou, du moins, les circulations d'eaux océaniques sont limitées. Les eaux s'appauvrissent ainsi en oxygène ce qui peut créer des zones d'anoxie bassinales. Mais le phénomène d'anoxie plus généralisée, appelé événement anoxique océanique (EAO), parait plutôt se développer en début de déglaciation lorsque les contrastes de température et de salinité entre les eaux de dégel et les eaux chaudes des basses latitudes conduisent à une stratification des eaux océaniques responsable de l’anoxie et de la sédimentation d’argiles noires sur de vastes étendues du globe. Ces environnements anoxiques sont quasiment dépourvus de faune.
- Les teneurs estimées en dioxyde de carbone (CO2) dans les sédiments déposés lors de la glaciation de la fin de l'Ordovicien sont élevées, ce qui apparait paradoxal car de fortes teneurs en CO2 sont caractéristiques des périodes chaudes de la Terre dites de "super effet de serre" (Greenhouse earth ou hothouse en anglais). Ces fortes teneurs sont en fait une conséquence de la glaciation : l'extension des calottes polaires empêche l'altération des silicates (phénomène qui est un grand consommateur de CO2 par la réaction : silicates + CO2 + H2O à cations + bicarbonate + SiO2). Cette augmentation des teneurs en dioxyde de carbone va se traduire assez rapidement par une élévation des températures et le début de la déglaciation.
- L'impact sur le monde vivant est considérable avec deux pics principaux d'extinction : un premier en début de glaciation, provoqué par la chute assez rapide du niveau marin et un second à la fin de la glaciation lorsque le niveau marin remonte et que les animaux marins doivent se réadapter à de nouveaux habitats.
- L'extinction de l'Ordovicien-Silurien aboutit à la disparition de 27% des familles et de 57% des genres d'animaux marins. Elle est considérée comme la seconde plus importante des cinq grandes extinctions massives du Phanérozoïque après celle du Permien-Trias qui interviendra environ 200 millions d'années plus tard.
- Parmi les groupes d'animaux marins fortement affectés par cette phase d'extinction :
- Au sein du plancton: les chitinozoaires et les acritarches,
- Les graptolites,
- Les trilobites,
- Les brachiopodes,
- Les coraux tabulés et les coraux tétracoralliaires,
Les conodontes, etc.
Le taux de disparition de familles d'animaux marins dans la partie supérieure de l'Ordovicien, sur environ 20 millions d'années est le plus élevé jamais enregistré au cours de l'histoire de la Terre, il est de l'ordre de 20 familles par million d'années.
Les premières communautés terrestres : des écosystèmes dynamiques>
Les premières traces de colonisation du domaine terrestre par des organismes complexes datent de l'Ordovicien inférieur. Elles sont rares mais permettent de reconstituer les principales étapes de cette colonisation.
Une question doit cependant être débattue au préalable : pourquoi à l'Ordovicien, pas plus tôt, pas plus tard ? On peut, avec beaucoup de prudence, proposer quelques pistes.
Le Cambrien et l'Ordovicien sont des périodes d'intense activité tectonique : le continent Rhodinia se fragmente et le Gondwana se forme par collision de ses parties Est et Ouest. Cette intense activité est accompagnée d'importantes variations du niveau des océans : à la transgression du Cambrien (transgression glacio-eustatique due pour partie à l'activité tectonique, pour partie à la fin de la glaciation du Vendien) qui étend sur les continents d'immenses mers épicontinentales peu profondes, suit un "aller-retour" marin à la fin de l'Ordovicien (régression puis transgression de l'Ashgill).
Cette dernière oscillation marine est directement en relation avec l'importante crise que subit la biosphère à la fin de l'Ordovicien et connue sous le nom de crise de l'Ashgill.
Les travaux sur les écosystèmes marins du Cambrien et de l'Ordovicien (de Burgess aux dernières découvertes en Chine) montrent qu'ils présentent une importante biodiversité : tous les grands groupes d'organismes sont en place, ainsi que bien d'autres qui ont ensuite disparu. Ces écosystèmes marins sont d'excellents réservoirs pour la colonisation continentale.
Les importantes modifications de leur environnement et de leur répartition (conquête des niches écologiques produites par la transgression, sélection drastique et brutale lors de la crise de l'Ashgill) ont certainement sélectionné des organismes aptes à partir à la conquête des continents par l'intermédiaire des estuaires et des marais salés (on sait qu'aujourd'hui ces milieux sont les portes d'entrée du domaine continental, aquatique comme terrestre, pour les colonisateurs marins). Cette conquête qui semble débuter à l'Ordovicien moyen, s'est certainement déroulée par vagues successives.
Les pressions sélectives liées aux modifications de l'environnement marin ne peuvent cependant pas être seules responsables de la colonisation du domaine terrestre : encore faut-il que ce domaine s'y prête !
Les études paléo-climatiques montrent que la teneur en CO2 atmosphérique est importante au Cambrien et à l'Ordovicien inférieur (18 fois plus qu'aujourd'hui) : l'effet de serre intense qui en résulte provoque des températures moyennes d'environ 40°C en été, avec des climats arides ou semi-arides au niveau de l'équateur. Des conditions peu favorables à la sortie des eaux.
Cependant le climat change au cours de l'Ordovicien supérieur : l'effet de serre semble diminuer (en partie par enfouissement important de carbone organique produit par un plancton extrêmement actif, en partie par baisse de l'activité volcanique) alors qu'un gradient de températures se remet en place entre l'équateur et le pôle Sud, permettant le développement d'une calotte polaire Sud.
Ainsi, certaines régions seraient devenues favorables à la sortie des eaux en offrant un climat humide et doux.
Autre paramètre rendant les terres émergées accueillantes aux premiers végétaux : la présence de sols mettant à disposition des premiers végétaux des éléments minéraux mobilisables et une surface d'ancrage pour leurs rhizoïdes.
La formation de paléosols est en partie due aux bactéries, cyanobactéries mais aussi aux lichens dont les premières traces fossiles datent de 400 Ma : leur activité biogéochimique aurait été favorisée par le haut niveau du CO2 atmosphérique (augmentation de l'activité photosynthétique et acidification des pluies qui décomposent plus facilement les sols).
Précambrien Les cyanobactéries du domaine intertidal, notamment, sont adaptées à des variations
importantes des conditions environnementales (alternance de conditions sèches et
humides, fluctuations de la salinité).Les premiers organismes à coloniser les surfaces continentales furent certainement des
cyanobactéries : encroûtements bactériens fossilisés et paléosols enrichis en matière
organique.Cambrien moyen Embryophyte fossile classé parmi les Bryophytes (Kaili Formation, Chine) Plus vieilles traces de cryptospores trouvées avec des fragments de tissus sporogènes
(Kaili Formation, Chine)Ordovicien inférieur Cryptospores datent de la fin de l'Ordovicien inférieur (-475 Ma) d'Arabie Saoudite Les cryptospores sont des spores d'affinité incertaine mais
considérées comme produites par des végétaux terrestres.Nombreuses traces de la présence potentielle d'organismes végétaux et animaux : des paléosols
dont l'enrichissement en matière organique pourrait porter la signature chimique de végétaux
terrestres (Juniata Formation, Pennsylvanie, USAPremiers champignons ascomycètes (dépôts du Ludlovien de Suède). Leur organisation fréquente en tétrades en fait
des méiosporesOrdovicien supérieur Les cryptospores ordoviciennes ont été identifiées un peu partout dans le monde :
la flore qui les a produites était donc certainement universelle.Spores découvertes dans des schistes du Sultanat d'Oman et vieux d'environ 450 Ma (Caradoc). De par leur organisation en tétrades (suggérant qu'elles sont issues de méiose, donc produites par
des sporophytes), leur taille et la structure de leur paroi (imprégnée de sporopollenine, polymère
complexe fournissant une protection contre la dessiccation et les UV) ainsi que leur organisation
au sein de restants de sporanges, ces cryptospores sont attribuées (par actualisme mais aussi par
comparaison avec les flores du Silurien et du Dévonien) à des Embryophytes, certainement
proches des actuelles hépathiques (marchantiophytes).Silurien. Premiers fragments de cuticule et premières cellules conductrices de type trachéide (la présence
de lignine, considérée comme possible, n'a pu être prouvée) dès le Silurien inférieur (- 430 Ma).
Cuticule et tissus conducteurs sont deux adaptations importantes à la vie terrestre des
végétaux : leur absence des dépôts plus anciens peut s'expliquer par leur fragilité
vis-à-vis des modes d'extraction mais aussi par leur fragilité vis-à-vis de la conservation. On
pense notamment que la nature chimique des composants de la cuticule facilite sa destruction
avant fossilisation. Les stomates n'apparaissent que plus tard (- 408 Ma).On peut considérer que les végétaux colonisent les terres
émergées au plus tard à l'Ordovicien moyen : ce sont les
premiers Embryophytes (sporopollenine + cuticule).Leurs adaptations aux conditions terrestres, en partie
héritées de leurs ancêtres aquatiques, leur permettent de
se répandre (spores dispersées par le vent) et se diversifier
sur des domaines alors vierges : les trachéophytes (tissu
conducteur constitué de trachéides) sont présents dès la
base du Silurien.Un paléosol du Ludlovien (fin du Silurien) de Pennsylvanie montre des traces ressemblant à celles
laissées par des rhizomes, indiquant la possible présence de trachéophytes.De possibles pelotes fécales sont associées à des hyphes d'ascomycètes trouvés dans des
dépôts du Ludlowien (fin Silurien), suggérant la présence de microarthropodes mycophages.Les animaux ne sont identifiés avec certitude qu'à la fin du
Silurien, suggérant une colonisation retardée par rapport
aux végétaux ou une colonisation par des organismes
à corps mou !La colonisation des terres émergées est réalisée, à l'Ordovicien, dans un contexte global favorable : biodiversité marine importante au sein d'écosystèmes en pleine évolution, interactions océan-continent importantes (transgressions-régressions, mers épicontinentales), climats diversifiés.
Les Embryophytes, certainement apparus en milieu marin au Cambrien, colonisent les terres émergées à l'Ordovicien supérieur (peut-être inférieur). Certaines structures et fonctions, mises en place en milieu marin, peuvent avoir été détournées de leur rôle premier (exaptations) et faciliter l'adaptation aux conditions de vie en milieu aérien.
Les données récentes en provenance de Chine (site de Kaili) ainsi que les diverses données anciennes sur les spores, indiquent cependant que cette colonisation a commencé bien plus tôt, au Cambrien. Cette donnée confirme certaines analyses génétiques qui placent l'apparition des Embryophytes au Cambrien, voir dès le Précambrien.
Le taxon des Embryophytes regroupe l'ensemble des végétaux terrestres, des anciennes mousses aux Angiospermes. Les données fossiles comme certains travaux à l'aide d'horloges moléculaires placent l'ancêtre commun des Embryophytes au début du Cambrien.
L'hypothèse aujourd'hui privilégiée quant à l'origine des Embryophytes est qu'ils ont évolué à partir de chlorophytes (anciennes algues vertes), certainement d'eau douce, avec lesquelles ils partagent nombre de caractéristiques ultra structurales, biochimiques, génétiques et morphologiques. Hypothèse développée en 1890, on peut aujourd'hui sans ambiguïté assurer que les Embryophytes sont des Chlorobiontes (groupe monophylétique).
Des études plus poussées montrent qu'il existe une proximité importante, au sein des chlorobiontes, avec les Charophytes avec lesquelles elles sont d'ailleurs désormais regroupées, dans le taxon des Plasmodesmophytes.
Les Charophytes sont des algues d'eau douce qui présentent des caractères dérivés communs avec les Embryophytes : l'enzyme Cu/Zn super oxyde dismutase (qui protège les cellules végétales des dommages occasionnés par les radicaux libres issus d'O2 ) et la sporopollenine, considérées comme des adaptations à la vie terrestre, sont également présentes chez ces algues d'eau douce. Leur présence chez l'ancêtre commun des deux groupes aurait donc pu faciliter la conquête du milieu terrestre par des Embryophytes aventureuses !
Ce site écossais d'une richesse paléontologique exceptionnelle a été décrit et cartographié pour la première fois en 1878. Depuis, de très nombreux travaux y ont été réalisés dans tous les domaines de la géologie, de la lithostratigraphie à la reconstitution des paléo-environnements locaux, en passant bien sûr par la paléontologie et la paléoécologie.
Continent Laurussia composé de l'Amérique du Nord, du Groenland, de l'écosse, de l'Angleterre et du Pays de Galles et portant le site fossilifère de Rhynie.
www.abdn.ac.uk/rhynie
Carte géologique simplifiée du site de Rhynie.
www.abdn.ac.uk/rhynieLes cherts fossilifères ont 396 +/- 8 millions d'années, d'après l'analyse radio-chronologique réalisée à partir des isotopes 39 et 40 de l'argon : ils appartiennent donc à la fin du Dévonien inférieur (Emsien).
L'origine de cette roche sédimentaire (roche siliceuse formées essentiellement de calcédoine et / ou d'opale et produite par des sources chaudes, nombreuses en raison des activités volcaniques de la région) et donc les conditions de fossilisation qui lui sont généralement associées font de Rhynie un site exceptionnel car :
1. Les organismes vivants y ont été conservés dans des conditions rares. Végétaux et animaux peuvent y être étudiés pour certains, jusque dans leurs ultra structures cellulaires.
2. Les communautés ont pu en partie être reconstituées dans leur diversité et dans leurs interactions.
Les conditions de fossilisation à Rhynie, il y a 396 millions d'années.
www.abdn.ac.uk/rhynieLe site de Rhynie a conservé une étape importante de la diversification des premiers organismes terrestres : on y a retrouvé huit genres de végétaux !
La flore de Rhynie comporte aussi des algues d'eau douce (dont des Charophytes), des champignons (les premiers Ascomycètes identifiés avec certitude et des Chytridiomycètes engagés dans des relations écologiques préservées) mais aussi le premier "lichen" identifié.
La préservation de la diversité et des particularités (anatomiques et écologiques) des organismes vivants a permis de reconstituer le premier écosystème complet de l'histoire de la vie en domaine terrestre.



Les Embryophytes du Dévonien inférieur de Rhynie.
De gauche à droite : Aglaophyton, Horneophyton, Nothia, Rhynia.

De gauche à droite : Asteroxylon et Ventarura.

A gauche : Zoosterophyllum (taxon des Lycophytes) (A : milieu de vie aquatique, B : sporanges réniformes en paires alternées au sommet des axes, C : reconstitution d'ensemble avec rhizome ramifié. A droite : Psilophyton sp (taxon des Euphyllophytes) reconstitution d'ensemble montrant le rhizome, les sporanges terminaux, des axes fertiles et les microphylles épineuses.
Dessins www.bio.indiana.edu/ , www.bio.ilstu.edu/ , www.ujf-grenoble.fr/choler/BEV/cour , Noëlla Wébert et L.V.B.
Paysage ordovicien reconstitué. Cette reconstitution montre le lien encore très fort avec le milieu aquatique dont les végétaux sont issus, comme les sites contemporains dans un domaine marécageux.
https://www.lepoint.fr/environnement/cretace-permien- devonien-les-principales-extinctions-de-masse-29-04-2019-2309993_1927.phpLes végétaux présents possèdent cependant les adaptations au milieu terrestre apparues dès l'ordovicien supérieur ou le Silurien inférieur, telles que la cuticule et les stomates. Et pour certains un tissu conducteur composé de trachéides. On note également la répartition des végétaux en "îlots" monospécifiques. Cette répartition est due à la croissance de nombre d'espèces à l'aide de rhizomes (rhizomes fossilisés ou présents sur des fossiles du même genre trouvés dans d'autres sites ordoviciano-siluro-dévonien) : les spécialistes considèrent, par application du principe d'actualisme, que ces îlots sont construits par des clones, au sein desquels la reproduction sexuée est rare.
Les milieux comme celui de Rhynie, ouverts et humides, présentent une importante disponibilité en ressources (lumière, eau, éléments minéraux, mais pas encore d'humus car la production de litière par ces communautés basses et peu feuillues était certainement limitée) : la comparaison avec des milieux équivalents (sommets de vastes blocs rocheux, bas-fonds, abords de grandes cascades) actuels montre que les espèces recouvrent rapidement ces milieux par multiplication végétative préférentiellement. Ce mode de croissance des organismes permet une occupation rapide de l'espace disponible sous l'effet d'une compétition interspécifique (mais aussi intraspécifique puisque les îlots sont des clones) importante : une stratégie adaptée pour des organismes dont les capacités d'exploitation du milieu sont limitées, tant pour l'approvisionnement en eau et sels minéraux (pas encore de racines vraies) que pour la capture de lumière (certaines espèces possèdent seulement des microphylles, les axes des autres sont nus).
Dans les milieux actuels, cette compétition limite la biodiversité : on peut donc supposer que le début de la colonisation du milieu terrestre s'est faite assez rapidement, sous l'effet d'une importante compétition entre les espèces végétales peu nombreuses (un peu suivant un modèle "tête de pont"), la diversification ne s'étant réalisée qu'après, dans des milieux déjà modelés (par exemple les sous-bois du Dévonien supérieur). La répartition des fossiles dans les dépôts montre qu'ils peuvent également se succéder sur un site lorsque celui-ci est modifié : les végétaux de Rhynie présenteraient une certaine spécificité écologique. Ainsi, le genre Rhynia semble être un colonisateur précoce de sols drainés à litière végétale pauvre, que l'on trouve d'ailleurs plutôt en dépôts monospécifiques. Par contre, les genres Asteroxylon et Aglaophyton seraient plutôt associés à des sols assez secs, riches en matières organiques et présents en groupements diversifiés, comme Trichopherophyton, colonisateur tardif de sols riches.
Cette succession écologique probable montre l'impact des végétaux sur l'évolution des sols qu'ils stabilisent et enrichissent en matière organique (formation d'une litière même si la production de biomasse végétale est faible). Les végétaux vont également modifier la configuration réseau hydrographique et en limiter les débordements tout en drainant des surfaces toujours plus importantes, qui offriront ainsi de nouvelles niches écologiques. L'apparente simplicité de ces organismes ne doit pas tromper : la diversité des adaptations écologiques est associée à l'évolution anatomique, bien marquée à l'Emsien. Tout d'abord, tous ces végétaux présentent des rhizoïdes fixés à leurs rhizomes ce qui rend l'eau et les éléments minéraux du sous-sol exploitables. Les végétaux peuvent coloniser des terrains un peu plus secs (du moins en surface) qu'auparavant. Ensuite, certains de ces végétaux (au moins cinq genres à Rhynie) sont vasculaires : ils présentent un xylème contenant des trachéides. Ce transport de l'eau dans des cellules aux parois rigidifiées est indissociable d'une croissance verticale nécessaire à la capture de la lumière.
Les Embryophytes de Rhynie présentent d'ailleurs toutes une croissance verticale affirmée : leur taille dépasse 20 cm de haut, certains développent un port buissonnant (jusque 1 mètre de haut). La capture de la lumière est facilitée par le développement, chez certains végétaux (Asteroxylon), des premières "feuilles", les microphylles. Rhynie montre donc des végétaux qui se partagent l'espace et les ressources, qui se répartissent (dans le temps et dans l'espace) en fonction des conditions du milieu et de la compétition interspécifique, qui modifient le milieu dans lequel ils se développent, de la colonisation de domaines marécageux originels à la structuration de véritables sols et la modification de l'environnement physique (réseau hydrographique, échanges gazeux et minéraux).
Rhynie nous présente une véritable communauté végétale en pleine évolution. Mais les Embryophytes ne sont cependant pas seuls à Rhynie. La place prise par les champignons, difficile à apprécier à sa juste valeur tant la fossilisation de ces organismes est difficile et aléatoire, n'en semble pas moins importante. Présents dans la litière fossilisée, ils sont également associés à d'autres organismes dans des relations symbiotiques (le "pseudo-lichen" Winfrenatia reticulata, dans lequel le photobionte est une cyanobactérie mais dont l'organisation est inconnue dans les lichens actuels) ou de parasitisme (parasitisme sur une charophyte, Palaeonitella cranii qui montre des réactions d'hypertrophie tissulaire dans les secteurs où les hyphes sont présentes, mais aussi mycoparasitisme d'un chytridiomycète sur un autre champignon).
L'évolution des communautés d'Embryophytes ne semble donc pas seulement soumise aux fluctuations du milieu ou à la compétition entre organismes proches, mais à une diversité d'interactions que l'on retrouve aujourd'hui dans tous les écosystèmes, même les plus simples. Ces interactions entre différents groupes d'organismes vivants confortent la notion de communautés colonisatrices. Les relations des Embryophytes avec leur environnement ont donc un aspect résolument moderne, en partie parce que cet environnement doit être entendu au sens large, incluant les facteurs édaphiques, hydrogéologiques et climatiques, mais aussi biotiques (compétition, parasitisme, symbioses).
Quelle place occupent les animaux à Rhynie ?
Les seuls animaux fossilisés sur ce site sont des arthropodes, ce qui n'est pas en soi même étonnant : malgré l'apparition précoce des "vers" la fossilisation de leurs tissus mous est difficile. Quant aux gastéropodes terrestres, leur apparition semble plus tardive. On trouve à Rhynie des arthropodes prédateurs (des Trigonotarbides et des myriapodes armés de forcipules) et des détritivores (acariens, le myriapode Leverhulmia mariae découvert avec le tube digestif rempli de spores et de débris végétaux, mais aussi le collembole Rhyniella praecursor, premier hexapode connu). L'examen approfondi d'un fossile resté mystérieux, Rhyniognatha hirsti , a amené M. Engel (Université du Kansas) et D. Grimaldi (AMNH de New York) à l'identifier comme le premier insecte, peut être très proche des Ptérygotes (insectes volants) avec lesquels il semble partager des caractères dérivés. Par contre, son statut de prédateur ne semble pas avoir été rediscuté.
Cette rapide présentation de la faune de Rhynie, diversifiée (par rapport aux sites fossiles plus âgés) présente cependant une lacune : il n'y a pas d'herbivores ! On ne trouve pas de trace conservée sur les végétaux d'attaque par mandibules, de découpage d'organes à l'état vivant : toute la matière végétale consommée semble être morte, certainement déjà modifiée par l'action des champignons et des bactéries. Il existe un doute sur des marques portées par un axe vertical de Rhyniophyte ressemblant à une piqûre, peut être laissée par un suceur de sève.
A Rhynie, il semble donc que le transfert de la matière végétale vers les consommateurs ne se fait pas par les consommateurs primaires que sont les herbivores mais par les détritivores. Cette organisation particulière du réseau trophique n'est cependant pas si extraordinaire qu'il y paraît : on l'observe actuellement dans la litière où les herbivores sont très rares ou absents. Ce qui est inattendu est que cette organisation sans herbivores s'observe dans un "macro-écosystème".
Les animaux profitent du gîte et des déchets plus que du couvert ! Ils s'installent dans un microenvironnement aux paramètres (hygrométrie, température,...) adaptés à leurs besoins et leurs caractéristiques physiologiques, et colonisent ainsi le domaine continental.
On peut alors penser que la présence des animaux n'a pas eu d'influence sur la dynamique évolutive des végétaux et de l'écosystème qu'ils ont construit. La réalité est certainement un peu plus complexe. Ainsi, les sporanges terminaux de la majorité des Embryophytes du Dévonien facilitent la dispersion des spores par le vent. Les animaux, cependant, auraient pu participer à cette dispersion, créant ainsi une première interaction animaux-végétaux non dénuée d'influence sur la dynamique des jeunes écosystèmes terrestres.
En quoi Rhynie est-il un écosystème particulièrement intéressant ?
Quatre points importants peuvent être soulevés :
1. Il est très bien conservé et propose une diversité des organismes, des situations et des relations, ce qui permet de reconstituer un paléo environnement complet dans lequel on peut comprendre le fonctionnement d'un écosystème dans son ensemble.
2. Il montre une étape importante dans l'évolution des végétaux de la Laurasia. On observe sur le même site une flore diversifiée, présentant des niveaux évolutifs différents (trachéophytes et non trachéophytes) organisés en une communauté végétale dynamique dans l'espace (colonisation du milieu en îlots monospécifiques) et dans le temps (succession d'organismes en interaction avec les modifications du milieu). Rhynie permet de comprendre les contraintes imposées aux Embryophytes par le milieu terrestre et leurs manières d'y répondre.
3. Il montre la diversité précoce des interactions écologiques qui animent les écosystèmes actuels : compétition inter et intraspécifique, parasitisme, symbiose...
4. Il montre que ces premiers écosystèmes possèdent un réseau trophique particulier, dépourvu des transformateurs habituels de la production primaire, les herbivores, et donc d'interactions directes animaux / végétaux. C'est l'évolution des plantes qui conduit l'évolution rapide des écosystèmes du Dévonien inférieur.L'écosystème découvert à Rhynie est-il représentatif des premiers écosystèmes terrestres, du milieu du Silurien au milieu du Dévonien ?
Les premiers macrofossiles d'Embryophytes identifiés sont classés dans le genre Cooksonia : les plus anciens proviennent de dépôts irlandais (Tipperary, 428 millions d'années, milieu du Silurien) et gallois. On en a découvert un peu partout dans le monde.
https://www.wikiwand.com/en/Cooksonia et https://steurh.home.xs4all.nl/eng/cook.htm
L'aspect très simple de cet organisme est trompeur. Comme le montrent le dessin et le fossile, le sporophyte de Cooksonia est ramifié (chaque ramification se terminant par un sporange) ce qui le classe dans la clade des Polysporangiophytes : cet organisme est donc plus évolué que les Bryophytes. Au centre de la tige, on observe dans certains cas un tissu conducteur composé de cellules ressemblant à des trachéides, classant Cooksonia dans le clade des Trachéophytes (végétaux possédant un tissu conducteur construit par des trachéides). Cooksonia est donc plus adaptée au milieu aérien que les Bryophytes chez qui le transfert de l'eau dans la plante se fait par des cellules hydroïdes qui ne sont pas organisées en un tissu différencié. Cependant, on a déterminé une certaine diversité parmi les fossiles découverts (forme des sporanges, présence ou pas de stomates) et le tissu conducteur central n'est pas toujours présent.
Cette diversité a incité certains paléobotanistes à déterminer quatre espèces de Cooksonia : le genre (ainsi que des genres proches et contemporains comme Renalai du Dévonien inférieur du Québec) serait plutôt un grade (un niveau évolutif) regroupant des organismes appartenant à des clades différents de Trachéophytes : les Rhyniophytes et les Lycophytes. D'autres auteurs les considèrent tous comme des Lycophytes. L'étude du genre Cooksonia montre que, malgré l'âge et les difficultés de conservation, on peut classer ces premiers fossiles dans des groupes végétaux identifiés à l'aide de caractères dérivés des végétaux terrestres : adaptation au milieu terrestre et diversification taxonomique semblent donc déjà bien engagés au Silurien moyen.
Il est plus difficile de reconstituer les environnements et les communautés associés à ces organismes : leur anatomie et les dépôts dans lesquels ils sont trouvés indiquent des communautés de vallées fluviales. Leurs axes, terminés par des sporanges, ont une croissance limitée : leur taille ne devait pas dépasser 10 cm. La production de litière était donc limitée, d'autant que les axes verticaux de Cooksonia étaient nus. Certains auteurs en ont déduit des communautés monospécifiques, organisées en îlots, à croissance courte permettant une production rapide de spores et à extension horizontale importante. Si tel est le cas, l'organisation et la dynamique des communautés végétales, comme celle observée à Rhynie, est déjà ancienne : elles se seraient mises en place presque en même temps que les premiers colonisateurs !
Des premiers dépôts à Cooksonia jusqu'aux premières forêts du Dévonien moyen, les communautés végétales dominées par les Trachéophytes seront constituées de deux clades majeurs : les Rhyniophytes (à sporanges terminaux) et les Lycophytes (à sporanges latéraux et donc à croissance prolongée par rapport aux Rhyniophytes). Si les Rhyniophytes sont seuls représentés à Ludford Lane (Angleterre, fin du Silurien), les deux clades sont représentés à Rhynie (Dévonien inférieur). Dans des dépôts à peine plus anciens (base du Dévonien moyen, - 395 Ma), on identifie le premier Euphyllophyte connu, Psilophyton : au Dévonien moyen, tous les grands taxons actuels sont en place !
les Cooksonia sont de petite taille mais les genres composant la flore de Rhynie dépassent les 18 cm. Le gain de taille est mis en relation avec une compétition importante pour la lumière, compétition dans laquelle le rôle de la feuille a certainement été déterminant. Alors que rien qui puisse ressembler à une feuille n'est identifié chez les Rhyniophytes, Asteroxylon porte de petites expansions latérales et subhorizontales appelées énations. A leur base, on observe la trace d'une petite expansion de la protostèle, considérée comme une ébauche de vascularisation. Chez le Lycophyte Leclercqia, du Dévonien moyen, l'énation est complètement vascularisée : on l'appelle microphylle. La microphylle, qui apparaît chez les Lycophytes est considéré comme le premier organe photosynthétique dans l'histoire des végétaux terrestres. Elle se généralise au cours du Dévonien moyen et supérieur, et on l'observe, par convergence, dans d'autres groupes contemporains (Sphénopsides = clade des prêles actuelles et des Calamites fossiles).
Les paléobotanistes pensent que l'énation est, à l'origine, une écaille protectrice des sporanges latéraux. Son évolution probable par extension et vascularisation fait de la microphylle une exaptation (une exaptation, concept défini par la paléontologue et évolutionniste américain Stephen Jay Gould, est une réutilisation opportuniste d'un caractère adaptatif liée à un changement de milieu et donc de pression de sélection). Elle n'est donc pas l'ancêtre de la feuille vraie (= mégaphylle) que l'on voit apparaître plus tard.
Les données provenant des différents sites de la future Laurasia (Euramérique, Sibérie, Chine) indiquent que la diversification des végétaux se poursuit tout au long de cette période chez les Lycophytes (premières Zoostérophylles au Gédinnien) et chez les Rhyniophytes. On identifie d'ailleurs, pour la future Laurasie (Euramérique, Chine, Sibérie) trois provinces biogéographiques dans lesquelles sont retrouvés nombre de genres communs. Certains genres (Zoosterophyllum) sont également présents dans les deux provinces gondwaniennes. Mais on trouve à l'est du Gondwana une province australienne présentant une flore particulière, identifiée dans l'état de Victoria, et dont la plante caractéristique est une Lycophyte, Baragwanathia longofolia. L'âge des dépôts dans lesquels elle a été trouvée a été l'objet de longues discussions et de nombreuses mesures. Un consensus semble s'établir sur la fin du Silurien (- 420 millions d'années). Si cette datation a été si débattue, c'est à cause de l'anatomie de cette plante qui présente des microphylles dans lesquelles on peut voir un vaisseau (non ramifié) ! De plus, les formes plus récentes possèdent des racines, et on suspecte donc la présence de racines chez les formes du Silurien. La comparaison avec les végétaux contemporains de l'hémisphère Nord (Cooksonia), bien plus primitifs, ou avec les Lycopsides présentant le même type d'organisation (Leclercqia ), bien plus récents, explique les nombreuses interrogations sur l'âge de Baragwanathia . Il apparaît difficile de la considérer, malgré son grand âge, comme une ancêtre chez les Lycophytes ! L'origine de ce groupe et le début de son histoire évolutive restent donc mal compris. On peut supposer que les Lycophytes ont évolué à des vitesses différentes dans les différentes régions de leur aire de répartition, celle du Gondwana acquérant très tôt des adaptations poussées à la vie terrestre.
L'origine monophylétique du groupe des Lycophytes ne semble, en effet, pas avoir été remis en question. Cette organisation en 5 provinces floristiques réalisée sur les données morphologiques comme sur les données taxonomiques montre qu'il existe un lien certain entre les traits morphologiques des végétaux siluro-dévoniens et les conditions climatiques. Par exemple, les sporanges regroupés (comme chez Psilophyton et Pertica) sont analysés comme des marqueurs de climats secs et/ou froids, le regroupement permettant de limiter leur dessiccation. L'influence des conditions environnementales apparaît donc important sur très jeunes communautés végétales et donc sur l'évolution et la diversification des premiers Embryophytes.
Comment se situe la faune de Rhynie par rapport à la dynamique des arthropodes du Siluro-Dévonien ?
On trouve des carnivores, des détritivores et des saprophages.
- Les arthropodes font leur entrée en force dans le Silurien supérieur (au moins en tant que fossiles identifiables au niveau des genres), ce qui indique que leur histoire est bien antérieure, certainement ordovicienne comme de nombreux éléments fossilisés (restes de cuticule, pelotes fécales et traces d'activités) le laissent penser.
- - De même, la diversité des premiers prédateurs à Ludford Lane (au moins deux groupes, les myriapodes scutigéromorphes et les trigonotarbides, qui ressemblent à des araignées) indique que des chaînes alimentaires étaient déjà en place à cette époque. Ces animaux chassaient donc des herbivores et/ou des détritivores.
- - Leur diversité augmente au cours du temps, tant au niveau des genres (pour les trigonotarbides, on trouve 9 espèces dans 3 genres à Gilboa contre 5 espèces dans deux genres à Rhynie et une seule espèce à Ludford Lane) qu'au niveau des groupes (presque tous les groupes d' Arachnides sont représentés à Gilboa, tous les groupes d'arthropodes terrestres sont apparus au Dévonien moyen).
- - Comme à Rhynie, aucun des autres sites connus de cette période ne livre d'herbivores identifiables avec certitude. Les arthropodes non carnivores sont tous (par actualisme ou par étude du contenu de leur tube digestif lorsque c'est possible) considérés comme des détritivores.
Cette organisation du monde animal va perdurer tout au long du Dévonien, et jusqu'à la fin du Carbonifère inférieur : on ne trouvera pas d'arthropode herbivore, même si leur présence est suspectée grâce à des arguments indirects (traces sur les feuilles, développement des tissus de protection sur certains organes végétaux).
Bilan sur les premier écosystèmes terrestres.
Dès le Cambrien (ou plus tôt encore ?), les Embryophytes conquièrent le domaine continental à partir des milieux humides (vallées fluviales, marais) connectés au domaine marin dont ils sont issus.
Face aux conditions difficiles du milieu aérien (dessiccation, difficulté d’une alimentation minérale régulière et suffisante, fixation de la lumière, dispersion des spores) ils vont développer, sous l’effet de la sélection naturelle, les adaptations nécessaires à leur implantation puis à leur dispersion (cuticule puis stomates, croissance verticale permise par le développement de tissus conducteurs lignifiés au sein d’une protostèle, microphylle, sporanges terminaux puis latéraux).
Certaines de ces adaptations (sporopollenine, cuticule) sont déjà présentes chez des végétaux aquatiques telles que les Charophytes et les Coleochaetales, groupes-frères des Embryophytes. Les Embryophytes les auraient donc affectées à de nouvelles tâches (exaptations ?). La rapide diversification des structures anatomiques peut-être mise en relation avec la rapide diversification phylogénétique dont les mécanismes et les étapes sont encore mal connus (origine des Lycophytes, relation entre les Rhyniophytes et les Lycophytes).
Au Dévonien moyen, les deux grands groupes d’Embryophytes "évolués" ou Trachéophytes, sont apparus : les Lycophytes (représentés actuellement par les prêles) et les Euphyllophytes, dominants aujourd’hui. Dès le Silurien moyen, cette conquête du domaine continental est le fait de communautés végétales, et non d’organismes isolés. Ces communautés sont écologiquement structurées et modifient de façon sensible leur milieu : formation de sols, drainage et stabilisation des surfaces conquises.
La dynamique évolutive des premiers Embryophytes et de leurs communautés semble avoir trois moteurs :
- La compétition intra et interspécifique (identifiée par le mode de croissance et de répartition des espèces) liée à la construction des communautés sur les domaines conquis.
- - Les symbioses (et surtout les mycorhizes).
- - L’influence des conditions environnementales.
Sur ce dernier point, la succession des espèces sur un site en fonction des modifications qu’il subit, bien étudiée à Rhynie, montre que les interactions environnement/végétal constituent un moteur important de la dynamique évolutive du taxon comme de la communauté végétale tout entière et l’organisation de la biosphère au Dévonien (au moins de son composant végétal) en 5 provinces floristiques confirme l’influence des conditions climatiques.
Il semble donc que, dans les premiers temps de la colonisation du domaine continental, les facteurs environnementaux aient eu une influence déterminante sur l’évolution et la distribution des végétaux. Les interactions végétaux-végétaux ont eu une influence déterminante au sein des communautés, stimulant la diversification au sein des provinces elles-mêmes. Par contre, l’absence d’animaux herbivores semble exclure une influence des interactions animaux/végétaux dans la construction et les premières étapes de l’évolution des écosystèmes terrestres.
Si on peut parler de communautés végétales, il apparaît plus difficile d’élargir ces communautés aux animaux : ceux-ci semblent plutôt se construire leur propre monde au dépend des déchets végétaux, certainement en partie métabolisés par les champignons et les bactéries, et à l’abri des groupements végétaux qui créent un microenvironnement favorable à la vie et à la reproduction des animaux. Au sein de ce monde animal, appendice commensale et phorétique des communautés végétales, les arthropodes se diversifient rapidement, au point que les arachnides (araignées, scorpions, acariens, amblypiges, pseudoscorpions) et les myriapodes (diplopodes et chilopodes) présentent l’essentiel des groupes actuellement connus, ainsi que quelques autres très importants à l’époque mais qui disparaîtront ultérieurement (trigonotarbides et arthropleurides qui disparaissent à la base du Permien).
Ces premières communautés organisent des écosystèmes apparemment découplés : des producteurs primaires et des consommateurs secondaires côte à côte, sans le lien établi dans les écosystèmes plus récents par les consommateurs primaires, et donc sans transfert direct de la production primaire du producteur au consommateur. Le lien est établi par les champignons et bactéries qui permettent aux détritivores (consommateurs secondaires dans ce cas) de se développer et de servir de proies aux différents prédateurs.
En réalité, les écosystèmes de la litière fonctionnent de la sorte puisque toutes leurs chaînes alimentaires sont basées sur la matière végétale (et animale) dégradée par les bactéries et champignons. Simplement, ces premiers écosystèmes de taille conséquente fonctionnent comme les écosystèmes actuels microscopiques.
Conclusion
A la fin du Dévonien moyen, les écosystèmes terrestres présentent une organisation, une dynamique évolutive et une biodiversité étonnantes au regard de leur jeune âge et des conditions que les organismes ont dû affronter en colonisant les terres émergées. Les transferts de matière et d’énergie, proches de ce que l’on observe actuellement dans la litière, ne permettent cependant pas, encore, un important développement de la biomasse : il manque dans ces écosystèmes le maillon essentiel de l’herbivore qui assure le transfert fondamentale de la matière végétale "fraîche" dans les chaînes alimentaires.
Revenons à "Brabantia", cette plaque qui s'est désolidarisée de Gondwana et qui comprenait le Sud de l'Irlande, le Sud de l'Angleterre, la Belgique, les Pays-Bas et le Nord-Ouest de l'Allemagne.
La région qui nous occupe (la Calestienne) est donc située au Sud-Est de ce micro continent en bordure de mer. Au large de cette région, toujours vers le Sud, on peut voir les falaises abruptes d'un haut plateau culminant à plus de 1000 m d'altitude avec des volcans parfois en activité (massif de Rocroi). Enfin, "Brabantia" se retrouve, il y a 385 millions d'années, au début du Dévonien, alors que la majorité des terres se situent à 30° de latitude Sud, réunie à "Baltia" (ou Baltica ) (la Scandinavie et la Russie et "Laurentia" (Amérique du Nord et Groenland) pour former un nouveau continent : "Laurussia" (ou Euramérica). Cette réunion produit le plissement calédonien qui fait émerger les terres, élève des chaînes de montagnes au Nord et produit un enfoncement de la partie Sud. Ces montagnes avaient des sommets qui culminaient à 1500 - 2000 m d'altitude et qui séparaient le plateau aride du désert du Nord des marais côtiers couverts de végétation luxuriante au Sud, région nous intéresse.
-410Ma Silurien Pridoli Pridolien
-415Ma Ludlowien Ludfordien Sortie de l'eau des plantes vasculaires Gorstien Actinoptérygiens -425Ma Wenlockien
Homérien Ostéichtyens, Myriapodes
-430Ma Scheinwoodien Gnathostomes Llandovérien Telychien -435Ma Aeronien Arachnides -440Ma Rhuddanien Sortie de l'eau des algues Le Silurien est un système géologique qui s'étend de -440 à -410Ma. La datation de début et de fin, bien que définie avec précision par les couches stratigraphiques de référence, est connue à seulement quelques millions d'années près. La fin de l'Ordovicien est marquée par une des cinq grandes extinctions massives de l'histoire des temps géologiques, où près de 60% des espèces marines ont disparu.
Une équipe de paléontologues chinois, américains et suédois a longuement "restauré" et étudié le fossile d’un poisson de 20cm appelé Entelognathus primordialis, découvert en 2010 dans la province du Yunnan (Chine) et vieux d’environ 420 millions d'années (Silurien supérieur). Il s’agit d’un placoderme, une classe éteinte de poissons cuirassés. Et - surprise ! - ce poisson, antérieur à la date vers laquelle la communauté scientifique situait généralement l’apparition des animaux à squelette osseux, possède déjà des mâchoires osseuses.
Ses os crâniens et ses maxillaires en font ainsi le plus ancien animal pourvu de mâchoires articulées. De quoi remettre en question une théorie largement admise, selon laquelle les Ostéichtyiens, un vaste groupe zoologique qui comprend tous les vertébrés à squelette osseux, ainsi que les poissons à squelette cartilagineux comme les raies et les requins, dérivaient tous de poissons à squelette lui aussi cartilagineux.
Dans ce modèle, requins et raies auraient conservé ce trait anatomique, tandis que d’autres poissons auraient évolué vers un squelette osseux (arêtes) puis engendré tous les tétrapodes (vertébrés terrestres, dont l’homme).
Or, le nouveau fossile, Entelognathus primordialis, pourrait être, sinon l’ancêtre direct de tous ces animaux, du moins l’un de ses proches cousins ou descendants. Le véritable ancêtre, lui aussi, devait donc forcément être déjà doté de ce trait observé chez Entelognathus et qui étonne tant ses découvreurs : une mâchoire osseuse. La lignée dont découle tous les vertébrés, osseux aussi bien que cartilagineux, possédait donc déjà un squelette osseux, estiment les auteurs de la nouvelle étude.
Le Silurien a été décrit par Roderick Murchison en 1839 (The Silurian System) se basant sur des terrains découverts en Galles du Sud. Il a été nommé en 1835 d'après une tribu celtique : les Silures.
Pendant le Silurien, le Gondwana reste dans les latitudes hautes de l'hémisphère Sud mais la calotte glaciaire est beaucoup moins étendue que pendant l'Ordovicien. La valse des continents suit son cours : le gigantesque archipel constitué des trois continents Sibéria, Baltica et Laurentia, situé au Nord du non moins immense supercontinent Protogondwana, se rapprochent les uns des autres pour peu à peu pour donner naissance tout au long du Silurien, à l'autre supercontinent du paléozoïque, le futur supercontinent Laurussia. Laurussia a donc été formé à la suite de la fermeture du grand océan Iapetus qui séparait les divers continents de l'hémisphère Nord avant leur accrétion. Ceci a donné naissance à l'orogenèse calédonienne, une chaine de montagnes qui traverse de part en part le jeune supercontinent et qui laisse entrevoir les prémices d’une autre chaîne de montagnes la future chaîne hercynienne qui se formera lorsque la Pangée émergera quelques dizaines de millions d'années plus tard au Carbonifère. On retrouve les traces de cette chaine de montagne en Scandinavie, au Canada et aux États-Unis où elle forme les Appalaches.
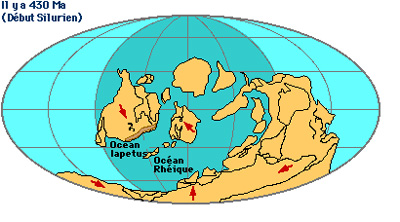
Au début du Silurien, il y a 430 millions d'années, Iapetus était devenu un océan étroit entre Laurentia et Baltica. Gondwana migrait toujours vers le Nord. L'espace océanique entre, au Nord Laurentia et Baltica, et au Sud Gondwana, a été appelé l'océan Rhéïque.
Au milieu du Silurien, il y a 420 millions d'années, l'océan Iapetus était presque refermé. Au Nord, la collision était imminente entre les deux plaques continentales Laurentia et Baltica.
Signalons ici qu'une grande partie de ce qui est aujourd'hui le Québec et l'Europe de l'Ouest se situent dans la zone tropicale au Sud de l'équateur. C'est pourquoi, par exemple, il s'est développé une grande barrière récifale qui va de la pointe de la Gaspésie, jusqu'aux Cantons de l'Est.
A la fin du Silurien, il y a 410 millions d'années, la collision Laurentia-Baltica a formé entre le Groenland et la Scandinavie la chaîne des Calédonides, une chaîne de montagnes qui est venue souder Baltica à Laurentia pour former une plus grande plaque continentale. Juste au Sud, le chaînon de microcontinents était sur le point d'entrer en collision avec la marge de Laurentia. A noter cet océan Rhéïque qui se referme progressivement entre Gondwana et le nouveau continent Laurentia-Baltica.
Les petits terranes accrétés au Gondwana pendant l'Ordovicien se détachent à nouveau pour former le superterrane hunnique, appelé ainsi car constitué de la majeure partie des territoires dévastés par Attila et les Huns. Les couches siluriennes ont produit du gaz et du pétrole dans certaines régions du globe tandis que d’autres couches de cette même période ont produit de l’hématite, un minerai de fer, qui a été important pour l'économie coloniale en Amérique du Nord.
L'océan autrefois florissant de vie durant la période ordovicienne est désormais moribond, suite à l'extinction massive survenue à la fin de l’Ordovicien, bon nombre de familles, de genres, d'espèces animales ont disparu. Mais la vie n'a pas terminé son combat pour autant. La nature ayant horreur du vide, dit-on, les espèces survivantes vont connaitre une radiation évolutive et combler les niches devenues vacantes.
La calotte glaciaire qui recouvrait le Gondwana à la fin de l'Ordovicien a maintenant presque fondu. Le niveau des océans s'est fortement élevé en Amérique du Nord-Est et en Europe. Les températures sont plutôt clémentes, tournant alentour des 20°C. Sur les terres, c'est encore plus ou moins la désolation. Des formes primitives de plantes multicellulaires envahissent les terres : quelques lichens et les premières mousses font leur apparition. Cependant, au bord de l'eau, là ou des sédiments ont été déposés par les fleuves, par les marées ou tout simplement par le flux et le reflux de la mer, certaines plantes peuvent prendre pied et s’accrocher. Ainsi des plantes comme Cooksonia, se développent arborant des tiges de quelques centimètres leur permettant de diffuser leurs spores dans les airs et ainsi se reproduire. Dans ce nouvel environnement, ces premières conquérantes de la terre ferme font face à la gravité, elles vont donc évoluer et se munir d’un "squelette végétal" et d'une "peau" protectrice les aidant à mieux supporter leur propre poids et la sècheresse. Et dès le Silurien moyen elles possèderont de minuscules orifices appelés "stomates" leur permettant une respiration aérienne.
D'autres plantes, un peu plus évoluées possèdent des cellules spécialisées, permettant le transport de l'eau et des nutriments vers les organes situés en hauteur. En outre, sur la terre, à l'écart du rivage, il n'y a pas ou peu de terre meuble et encore moins de nutriments, ce n’est quasiment que de la roche, elles vont donc développer des rhizomes, des sortes de racines-bulbes qui vont puiser le peu de nourriture présente dans le sol mais surtout la stocker. Plus tard, elles développeront des excroissances afin de capter plus de soleil, c'est l'apparition des premières feuilles. Ainsi commencera la course à la hauteur, qui finira par permettre aux futures fougères d’atteindre des tailles sans précédent.Les arthropodes étant très liés au règne végétal, même sous l’eau, on pense que la conquête des terres par cette première végétation, a permis à certains arthropodes de leur emboîter le pas. Par ailleurs, la mer rejetait toutes sortes de cadavres sur le littoral, une manne de nourriture très attrayante, ce qui en fait une autre raison probable de la sortie des arthropodes carnivores et charognards des eaux. Ainsi, il est probable que dès cette époque, de petits arthropodes et autres acariens peuplaient les berges des continents. Une chose est sûre : dès la fin du Silurien ou tout au début du Dévonien, ces petits animaux ont conquis le littoral ou prospèrent les mousses et autres végétations primitives, participant activement au recyclage de la matière organique, et à la formation des premiers sols. Mais cela reste encore aujourd'hui ouvert au débat.
Quoi qu'il en soit, dans les hauts fonds de l'océan Iapetus, bordant la Laurussia en formation et le Protogondwana plus au Sud, la faune marine est en pleine expansion. Outre la multitude d'animaux planctoniques qui explose, les coraux se multiplient et les premiers récifs font leur apparition, les crinoïdes s’épanouissent, les brachiopodes garnissent les fonds marins, les trilobites se diversifient, les nautiloïdes voient leurs formes de plus en plus variées, mais c'est surtout l'âge d'or des grands arthropodes marins, les fameux scorpions marins, les euryptérides. Ils sont devenus les plus grands arthropodes marins de tous les temps, certaines espèces atteignant la terrifiante taille de plus de deux mètres de long, tel le Ptérygotus. Ils sont, à ce moment de l'histoire les plus grands prédateurs des océans, et par extension, de la planète. Autant dire que les trilobites et autres bestioles benthiques n’en mènent pas large face à ces monstres marins.
Eurypterus est un genre éteint d'Eurypterid, un groupe d'organismes arthropodes communément appelés "scorpions de mer". Le genre a vécu pendant la période silurienne, il y a environ 432 à 418 millions d'années.
Mais la grande nouveauté, ce qui marque le Silurien, c’est l'apparition des premiers Placodermes. Si l'Ordovicien à connu les Ostracodermes, ces poissons en partie cuirassés et sans mâchoire, le Silurien voit apparaitre les poissons équipés de mâchoires, les Gnathostomes. Certes, il s'agit de mâchoires primitives, et bien que puissantes, les dents sont encore absentes. Néanmoins, la mâchoire des ces poissons ancestraux présentes des aspérités ou excroissances osseuses biens tranchantes faisant office de dents comme on peut le voir sur le fossile de Dunkléosteus, un Placoderme qui apparaitra au Dévonien. Quant au reste de leur corps, à l'image des Ostracodermes, celui-ci est recouvert en partie d'une épaisse cuirasse articulée protégeant la tête du poisson, et désormais ils possèdent des écailles sur la partie arrière de leur corps. En outre, à la différence des Ostracodermes, les Placodermes possèdent des nageoires dignes de ce nom. Ce qui fait de ces poissons de redoutables prédateurs, d'autant plus que plus tard au Dévonien, que certains d'entre-eux atteindront des tailles significatives, pouvant dépasser les 8 mètres de long.
Par la suite, les Gnathostomes, poissons à mâchoires, vont se diversifier et engendrer les deux grands groupes de poissons, d’une part, les poissons cartilagineux, les Chondrichtyens, et d’autre part, les Ostéichtyens, les poissons osseux. Les Chondrichtyens seront à la base des requins, raies et autres chimères. Les poissons osseux, engendreront à leur tour, deux grands groupes, les Actinoptérygiens, ou pour faire simple les poissons à nageoires rayonnée comme le mérou actuel, et les Sarcoptérygiens, poissons à nageoires charnues qui engendreront les Crossoptérygiens, tel le cœlacanthe actuel.
En résumé, le Silurien aura vu les prémices de la conquête de la terre ferme par les plantes et autres arthropodes, et l’apparition des premiers poissons primitifs. En gros, le Silurien pourrait être considéré comme une sorte de période de transition entre l’Ordovicien et le Dévonien.
Le Silurien affleure sur d'assez grandes surfaces en bordure méridionale du Massif du Brabant, depuis Lessines à l'Ouest jusqu'à la Mehaigne à l'Est. Son épaisseur, estimée à plus de 3,5 km et ses faciès turbiditiques à hémipélagiques, indiquent un domaine en forte subsidence dans un environnement de mer profonde.
On y reconnaît la suite de la troisième méga séquence avec les formations de Brûtia (shales foncés à fucoïdes, puis la coulée ignimbritique de l'eurite de Grand-Manil ou de Nivelles), Bois Grand-Père (schiste et siltite gris, minces bancs de grès), Fallais (siltites et schistes chloriteux verdâtres, turbidites), Corroy (turbidites plus grossières : grès et schistes gris), Petit-Roeulx, Steenkerke (schistes foncés), Froide Fontaine (schistes à intercalations gréseuses turbiditiques), Vissoul et Fumal (schistes à intercalations gréseuses : turbidites distales), Vichenet (schistes et siltites gréseuses localement carbonatées : turbidites) et Ronquières (turbidites de type Bouma). Ces formations constituent le Groupe de l'Orneau.
L'apport sédimentaire aurait un caractère centripète par rapport à la zone centrale du massif. Donc, dans les zones externes Sud du Massif du Brabant, les sédiments auraient une origine méridionale, comme l'indique l'allure des turbidites. Il en est de même dans la Bande du Condroz. La rapide augmentation de la subsidence dès la partie supérieure du Llandoverien suggère la formation d'un bassin d'avant-pays dans les zones externes du massif. La présence de faunes d'acritarches remaniés (Silurien à Lochkovien inférieur) dans le Dévonien Inférieur du bassin de Dinant atteste que la zone centrale du Massif du Brabant n'a pas été submergée avant le Ludlowien et que son démantèlement par érosion débute dès le Lochkovien. A l'Ouest du massif, la sédimentation conserve des caractères de plate-forme peu profonde de Renaix à Veurne.
Dans la région de Visé, un important édifice volcanique silurien est connu par sondage. La mer ne l'a submergé qu'au Dévonien Moyen, ce dont témoigne un conglomérat supposé Givétien. Cet édifice est constitué d'une masse métarhyolitique vitreuse à structure fluidale et de coulées de méta-obsidiennes.
Au Silurien, les massifs ardennais sont vraisemblablement en phase de plissement et n'enregistrent aucune sédimentation, excepté dans le Massif de Stavelot où la Formation de Petites Tailles aurait un âge pré-dévonien, peut-être Silurien supérieur (Pridolien).
Initialement, les formations de la Bande du Condroz devaient se localiser plusieurs dizaines de kilomètres plus au Sud que leur position actuelle par rapport au Massif du Brabant (puisqu'elles seront transportées ultérieurement par la Faille du Midi). Cependant, elles peuvent facilement se corréler stratigraphiquement avec les formations du Massif du Brabant ce qui permet de supposer la permanence du bassin unique Brabant-Condroz.
Pour me contacter, me faire part de vos idées, me poser vos questions, me laisser vos remarques, cliquez sur l'image ci-dessous...